| | 訂閱本報 | 瀏覽歷史報區 | | |
 |
|
| 【驚蟄】三月五日。驚蟄象徵春天即將來臨,萬物亦將活躍起來。 【春分】三月廿日。春分時天氣變化大,農作物較易發生病蟲害,田間防治工作不可少。 【諺語】驚蟄鳥仔曝翅:驚蟄後氣溫漸升,不僅土中蟲兒蠢蠢動起來,連鳥兒也振翅享受日光浴。 |
|
| 二〇一六年三月二十日發行 | |
米故事 臺灣稻米古典名種談(四) |
|
|
臺灣稻米古典名種談(四)
圖32 1922年後,西日本的霸主品種「旭」和東日本的霸主品種「亀ノ尾」(亀の尾),在「雜交育種法」中進行東、西血脈的大融合,構築了日本水稻雜交育種的「旭時代」。
圖33 育成「亀治」品種的篤農育種家:廣田 亀治(1839~1896)。 亀治(かめじ‧Kameji)與 亀ノ尾(亀の尾,かめのお‧Kamenoo) 「亀治」與「亀の尾」這兩個的「亀」字輩的品種育成時間相隔了23年,但它們卻有著極為嚴重的瑜亮情結,情結不是發生在品種本身之間,而是發生在兩位素昧平生、之後也未曾謀過面的篤農育種家。 「亀治」這個品種是由廣田 亀治在1875年(明治8年)育成的。日本明治政府是在1893年才開始設置國立農業試驗場,所以那時候絕大部分的作物育種的成就,都出自像廣田 亀治這一類很有實務經驗的「篤農育種家」之手,他們很喜歡用自己的名字來為品種命名,想把自己的名字流傳給後世。廣田 亀治是1839年(天保10年)6月5日出生在當時的出雲国能義郡荒島村(現在的島根県安来市荒島町)廣田家次男。父親廣田 平助是擔任當時松江藩的「郷蔵番」(ゴウグラバン)─鄉里地租米穀倉管理員;這是一種世襲制的工作。廣田 亀治長大後,從父親那裡繼承了這份職位,同時自己也耕種了28公畝的水稻田,他種的是地方品種:「縮張」。從1870年開始的三年間大熱天氣候不穩定,時而雨後艷陽,島根県地區的水稻田很不平靜了起來,病蟲害頻繁發生,尤其是稻農最畏懼的稻熱病也開始猖獗。專注於精進耕作技術,連婚姻大事都給耽擱,孓然獨身的廣田 亀治,就把28公畝田中的8公畝用來作試驗觀察,他在重複的年份裡,用現在的「純系選種法」針對稻熱病的耐病性進行選種,在1875年成功選到耐病性佳的品系,他就用自己的名字來加以命名。 廣田 亀治育成「亀治」是開日本水稻抗病蟲育種的先河,這個品種在日本從明治時代到昭和時代延續很久,主要就是它優異的耐病性,畿內以西的水田曾經是它的天下。日本近代史幾次有名的戰爭,像日俄戰爭、太平洋戰爭[二戰],導致日本糧食短缺的危機,「亀治」都扮演了很重要的餵養饑荒角色。人們有感於廣田 亀治雖只是一介農夫,卻能作出劃時代的貢獻,57歲英年孤單逝去,所以就在現在JR國鐵荒島站[荒島駅]附近的國道9號線的出口,建立了一座他篤農荷鋤的銅像,永遠紀念著他。 在臺灣,「亀治」並沒有出現在橋口 文蔵的三次引種名單內,它何時來臺已不可考,雖然它也是貴為明治時期的名種,卻一直沒有在臺灣早期的蓬萊米故事歷程中浮上檯面一展長才,一直到1920年代早期,在磯 永吉與末永 仁廣泛進行日本水稻品種間的雜交育種組合名單中才猛然出現,表現果然一鳴驚人,當初末永 仁用它做為母本,原始考量就是想借重它的稻熱病耐病性,顯然,「亀治」來到臺灣在產量與品質的表現不過爾爾,但是在育種家的口袋中,它始終沒有須臾間被遺忘過的。
圖34 人們有感於廣田 亀治雖只是一介篤農,卻能作出劃時代的貢獻,所以在現在JR國鐵荒島站附近的國道9號線的出口,建了一座他篤農荷鋤的銅像,永遠記念著他。
圖35 「亀治」:末永 仁1925年用它做為母本與「神力」雜交,原始考量就是想借重它的稻熱病耐病性。(農業試驗所檔案照片)
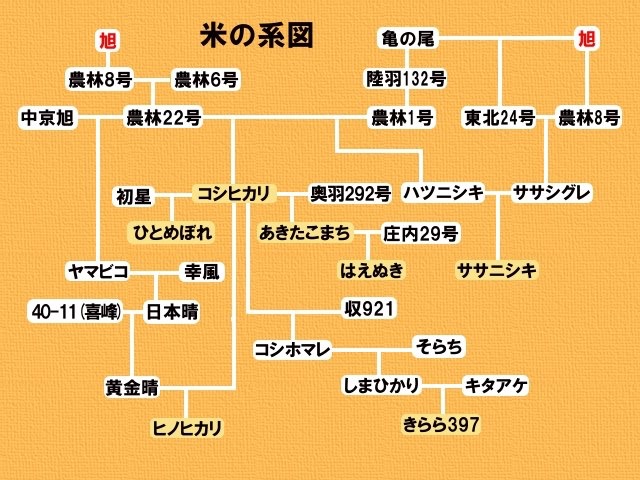
圖36 「亀の尾」:1920年代開始在日本水稻佔有一席之地,是東日本的霸主品種,在日本120年歷久不衰的頂級好米。 「亀の尾」是日本山形県東田川郡大和村篤農阿部 亀治(1868-1928)在1897年育成的。他所育成的「亀の尾」1920年代開始在日本水稻佔有一席之地,是東日本的霸主品種。它的「子」品種如陸羽132号(陸羽20号 x 亀の尾4号)、「孫」品種如農林1号(森多早生 x 陸羽132号)、「曾孫」品種如コシヒカリ(農林22号 x 農林1号)、「玄孫」品種如ササニシキ(ハツニシキ x ササシグレ);這些品種都是名列良質米之林,這也說明了為什麼吟釀業者會把「亀の尾」視同「夢幻的酒米」(幻の酒米)的緣由,後來日本有名的酒米品種有很多都帶有「亀の尾」的血緣。 阿部26歲(1893)那一年,是令稻農最擔心、心痛的冷害年。他前往位在立谷沢(現今的庄內町)的熊谷神社祭拜祈求老天保佑冷害快快過去,回程在路邊種植「惣兵衛早生」品種的水田中,看到三株寒風中屹立不搖的水稻植株,後來他就收了這三穗,這就是「亀の尾」的開始,四年後推出新品種。按那個年代的篤農育種家的傳統,他們喜歡用自己的名字來作為品種名,所以當初阿部 亀治在1897年推出新品種時,理當是名為「亀治」的;但是「亀治」之名早已在23年之前,已經被廣田 亀治搶先使用了,這個「亀治」就是在31年後,在臺中當起了「臺中65號」的母親。阿部的新品種,比起廣田 亀治的「亀治」實在是好太多了,他身邊的篤農朋友們很嚥不下「亀治」之名被人捷足先登這口氣,於是大家慫恿阿部將新品種命名為「亀王」(亀の王)〈讀音:カメノオウ / KamenoÓ〉以示抗衡,但是阿部極為無奈,又不好拂了眾人的美意,於是他「折衷」選了讀音很接近的「亀の尾」〈讀音:カメノオ / Kamenoo〉作為品種名。近年來,日本興起「活化」古典名種,尤其以「酒米」之姿重現江湖,「亀の尾」當然是首選不作第二人想的頂級米種;日本頗負盛名的漫畫與電視劇:《夏子の酒》[夏子的酒] 的腳本就是以新潟縣和島村的「久須美酒造」,用「亀の尾」釀造吟釀,推出吟釀品牌「亀の翁」〈讀音同「亀の王」〉的故事作為主所鋪陳的劇情;後來日本各家酒造,紛紛推出的品牌計有:「亀の翁」、「亀の王」及本尊「亀の尾」。所以「亀の尾」跟「旭」一樣,也是在日本120年歷久不衰的頂級好米。 「亀の尾」是1896年跟「旭」和「愛国」一起種在橋口 文蔵家門口那畝田[現在的北一女中]的第一批從日本引進的水稻品種,輩份是最高的。只是很可惜「亀の尾」在這畝田試作之後,就徹底從臺灣消失,它的表現完全跟它的同儕「旭」和「愛国」不能相比;它與「旭」曾經分別是「割據」東、西日本的霸主品種,它們同批在1896年來到臺灣,在臺灣蓬萊米的歷史裡,「旭」一直有它重要的角色與地位,但是貴為東日本霸主品種的「亀の尾」卻在1896年就一蹶不振,真讓人有「淮橘為枳」之嘆。
圖37 育成「亀の尾」品種的篤農育種家:阿部亀治(1868-1928)。
圖38 1927年(昭和2年)9月5日,阿部 亀治因育成「亀の尾」而獲日本政府頒授「藍綬褒章」表揚。同日,在日本山形県庄内町小出新田・八幡神社內,建了一座「阿部 亀治翁頌德碑」[農學博士橫井 時敬題字、書法家黑崎 (馨)研堂執筆],紀念阿部 亀治在這裏發現三株抗冷水稻因而育成「亀の尾」。
圖39 「神力」是一個稍晚熟品種,莖稈堅強比「程好」稍矮,以現在術語來說,他是一種「穗數型」的高產品種。 神力(しんりき‧Sinliki) 1877年(明治10年),住在近畿地方兵庫県揖保郡中島村63歲的篤農丸尾 重次郎,一如往常在自家的水田蒔下他最慣常種的兵庫県播磨町地方品種「程好」的秧苗。出穗後,他發現了三穗是無芒的。這就是「神力」品種的開始。 這三穗增殖採種過程時,丸尾發現它的穀種收量遠多於「程好」(ほどよし‧Hotoyosi);富於試驗精神的他,懷疑是他自家水田環境使然,所以他廣徵其他地方的農家參與「地方試作」,結果證實它能增產25%,興奮之餘,於是丸尾就將這個無芒的新選系取一個極為寫實、寫他的心情的名字:「器量良」(きりょうよし‧Kilioyosi)。這個品種後來表現實在極為出色,讓丸尾重次郎感覺肯定是有神明助力保佑,於是他又重新將它命名為「神力」。 「神力」是一個稍晚熟品種,莖稈比「程好」稍矮,但堅強,以現在術語來說,他是一種「穗數型」的高產品種,它的「短桿穗數多」的特性,剛好迎合日本魚肥和豆餅開始上市的時代,「神力」出色的耐肥性真的是搭了時代的順風車,從明治三十年代[1897~]致昭和初年[1926~],它一躍成為西日本全域的代表品種;大正8年(1919年)的最盛期耕種面積曾高達58萬公頃,全國水稻面積的20%都種植「神力」。這個西日本霸主品種的地位一直維持到昭和初年,才被新品種「旭」所取代。
圖40 「神力」:從明治三十年代[1897~]至昭和初年[1926~],一躍成為西日本全域的代表品種。(農業試驗所檔案照片) 「神力」之所能成為明治時代的「奇跡米」[明治のミラクルライス(miracle rice)],除了育種者丸尾 重次郎的運氣及慧眼識英雄之外,另一個率先建立採種制度的岩村 善六也是幕後大功臣。岩村 善六是兵庫県揖保郡余部村(現在的姬路市)人,當時他是河内村第二代村長,熱心公益,帶領村人整治揖保川水患,頗得人心;他一眼看出「神力」高產的特性,必須維持精純的優良穀種,方能長治久安。他登高一呼選擇績優農家組織「採種工會」,專責供應純良穀種。在還沒有公營的採種組織的時代,建立雛型的「原種·原原種體制」的他,功績不得不讓人另眼看待。 整個「神力」的育種、推廣過程,丸尾與岩村二人的「純系選種」、「地區試驗」、「原種·原原種體制」也都是後來1893年日本引進歐美農藝科技,建立農場試驗標準育種程序的必要流程。早年在那種農藝科學尚處混沌的時代,「神力」就有幸在這兩位有遠見的篤農所建立的制度,保持了歷久不衰的遺傳背景,不啻終致為獨霸西日本的當家品種,即便至昭和初年易地崛起的新品種「旭」,也是順著這套標準程序扶搖直上,瓜代了「神力」的昔日光采,進而雙棲「純系選種法」與「雜交育種法」成為西日本霸主。 然而,「神力」與「旭」這一雙前後任霸主品種的恩怨情仇,乃至瑜亮情結並沒隨情勢底定而稍有止息,反是繼續從日本延燒來到臺灣,讓它們仇人在異地狹路相逢再度糾結的就是橋口 文蔵。
圖41 藝術家畫筆下「神力」的著穗與粒型。 「神力」是橋口 文蔵規劃的第二批自日本農商務省農事試驗場九州支場引進的日本種水稻,同行來臺灣的都是一時之選,甚至有些還不乏是名列「明治年間名種」的其他九個成員:竹成、都、江戶、中村、穗增、巾著、三石、白玉、今長者,地位僅次於1896年第一批的亀ノ尾、愛國、旭。這些名種迢迢千里跋涉到臺灣,一起種在橋口 文蔵家門口那畝田[現在的北一女中],開始在全新的環境進行PK,讓人唏噓的是,看似再簡單不過的新種稻米生產試作,卻沒想到這些出身不凡的古典名種在這個新環境顛沛流離,每一個品種的命運大不相同,而對農場試驗頗具見地的引種主人卻也「出師未捷身先死」,更使臺灣蓬萊米的發展平添一頁心酸史。 1895年8月,橋口 文蔵在臺北文武町他家門前設立20公畝的日本水稻試作田,是臺灣農業農場試驗研究之肇始。1896年第一批引種試作之同時,跟他同為薩摩籓出身、又有親戚關係的第一任臺灣總督樺山 資紀[樺山 資紀生父姓橋口],在即將離職返國就任內務大臣新職的兩個月前,有感於橋口 文蔵留美學習美式大農場經營管理與農場試驗的專長,畢業返國後又在北海道開拓使歷練實務經驗,於是借重他出任臺北縣知事一職,管轄今臺北縣市、宜蘭及基隆區域的行政事務。橋口在北海道有豐富的處理原住民阿伊努民族[アイヌ民族]與日本人農墾新移民之間事務經驗,闢建特用作物專屬大農場村莊「留寿都村」與「壮瞥町大滝村」為人所稱道。臺北縣知事兩年[1896年4月1日-1898年5月3日]主要還是傾全力解決日本稻在管轄地區適地適性的栽培困頓。「神力」等第二批引種到達時間1899年,他已體弱多病卸下知事職務,第三批引種1900年,這些古典名種的試作也面臨從原來官邸門前20公畝水田,整併到臺北東門外新設立的臺北縣農事試驗場[今臺大醫院新館址][1901年改名為臺灣總督府臺北農事試驗場];1903年再遷至臺北廳大加蚋堡頂內埔庄[後為富田町329番地,就是現今的臺大農場及臺科大]時,橋口已在日本病逝,得年僅50歲。壯志雖未酬,但是他經手而來的日本水稻古典名種,特別在他曾經治理過的臺北縣境,找到「化外」之地繼續試作,「神力」也仍在行列之中接受著未知的命運考驗。
圖42 在日本興盛起活化古典名種的風氣,「神力」以「酒米」之姿重現江湖。 橋口 文蔵1896年出任臺北縣知事後,繼續掌管官邸門前那畝田,貫徹橋口 文蔵引種明治年古典名種雄心壯志與使命感的,就是臺灣總督府民政局殖產部農務課長,跟橋口 文蔵同為薩摩藩出身的押川 則吉(おしかわのりきち、1863~1918)。到後來,1897年押川返日就任新職以及1903年農事單位整併,這三批引種歸建在臺灣總督府臺北農事試驗場,這段期間可以說是這些品種最悲慘的時候。1907年臺北廳農會受總督府臺北農事試驗場之託,以「神力」領銜的五個品種,開始了所謂的「軍刀農政」走進臺北縣境內的農家進行試作,「神力」與「旭」這一雙後來在1920年代爭奪西日本霸主品種地位的恩怨情仇,就在此時也易地碰頭,此其時兩者尚在五五波的態勢,尤其在士林、板橋的模範田,受到抽穗期的困擾,幾乎全面慘敗;又三年,所有平地水田試作的敗象已經導致總督府的稻米政策定調在在來米的改良,然而臺北廳農會的專家卻在淡水、小基隆(三芝)、金包里(金山)各支廳轄下較低溫的「山手高地」(面山邊坡高地)一年一作的地方悄悄地找到了一線生機。再一年,這線希望擴充到頂雙溪(新北市雙溪)以及士林山邊水田;從此各廳農會群起效尤,紛紛從日本各縣、府引進更多的二線、三線品種加入試作,氣象為之一新。及至1918年,押川 則吉在日本因他案自縊身亡,這兩位當年率先引進試作的古典名種終於不負盛名鶴立雞群表現不凡,也印證了他們兩人先知的眼光,只不過歷經20餘年形勢大有轉變,浮出檯面的轉為「中村」領銜,1923年的「幼苗插植法」更使之定於一尊,「神力」與「旭」已落在其後了。也在此時,「旭」以後來居上之勢全面取代了原來「神力」在西日本的地位成為霸主。1929年,「中村」因為稻熱病為害嚴重,失去大臺北平原的主要耕種品種的地位,正式被「旭」所取代,「神力」再次輸給了「旭」,此恨綿綿可想而知。 然而,「旭」在1929年出頭,卻也在同年埋下被淘汰的肇因。就在這一年,臺中州立農事試驗場的育種家末永 仁用「雜交育種法」選出的「臺中65號」,以秋風掃落葉之姿,橫掃了1926年以「中村」為代表命名為「蓬萊米」及其以下的「內地種」日本稻的氣勢,成為第一個真正在臺灣本土育成的「蓬萊米」品種。「臺中65號」的父親就是「神力」─王子復仇記,為「神力」扳回在臺灣蓬萊米發展史的定位。 在紀錄中,這個大規模的雜交育種計劃起始於1922年;斯時也,總督府尚未解禁對日本稻的研究,被私下選定為雜交親本的有:伊予仙石、旭、晚生旭、愛國、改良愛國、神力、三井、龜治、相川、酒井金子、メ張、盤田朝日、大阪旭、丹後中稻、京錦、明治穗、與吉選、豐國、竹成、畿內中生;而其中「臺中65號」所來自的雜交組合─「龜治」 x 「神力」─絕對不是一個隨機的媒合。因為在末永 仁的紀錄裡,一定清楚有著日本愛知與畿內地區的農試單位早有這個組合,它所衍生的後代:愛知晚14號~16號、畿內晚60號、畿內中5、17、52、73號,這是讓育種家所不敢忽略親本的潛力,也是「神力」的魅力! 晚近,在日本興盛起活化古典名種的風氣,尤其是以「酒米」之姿重現江湖,沒有人敢或忘「神力」曾經有過的風華;於是,140年名種再見佳釀於肆,讓有識者為之陶醉不已,這在明治年間古典名種之林中幾「種」能夠?跨越時空競逐的浮浮沉沉,「神力」還會有一絲絲的怨懟嗎?
圖43 140年名種再見佳釀於肆,讓有識者為之陶醉不已,這在明治年間古典名種之林中幾「種」能夠? 謝兆樞 / 臺灣北部蓬萊米走廊推動聯盟‧磯 永吉小屋團隊 [ 待續 ]
|
|
 |
|
|
國內稻熱病相關研究 從病原菌生理分群到水稻抗病育種 水稻,為全世界的三大糧食之一,供養全球超過半數以上人口,尤其在亞洲地區更為主要的熱量來源。在臺灣,每年約有26萬公頃以上的稻作面積,為栽培面積最廣的作物,且為國人的主要糧食來源。然而,水稻栽培於田間必然受到溫度、降雨、日照、病蟲害等外在條件影響,在不利的生產條件下即可能造成產量減損或品質低劣的現象,尤其是全球暖化造成氣候的不穩定,高溫及強降雨的情勢加劇,更使得農民在生產時面臨巨大的挑戰。 稻熱病是水稻生產過程最常遭遇的病害之一,全球有超過85個國家有遭受危害紀錄,預估達到10~30%的減產。病原菌(Magnaporthe grisea)最適宜的生長溫度在25~28℃之間,配合濕度達90%以上即適合繁殖傳播,病原菌從分生孢子侵入水稻葉面細胞至重新繁殖產生新的分生孢子僅需4~5天的時間,因此一般在第一期作春雨或梅雨期間容易發生,且一旦稻株受到感染,疫情傳佈極為迅速。 稻熱病菌的生理分群 國內早在1958年前後,農業試驗所植物病理組簡錦忠博士即自臺灣各地蒐集菌株,並依據其對於16個水稻判別品種的抗感病情形,共區分為七群(P、I、J、T、K、N及A),78個生理小種,並由後續的監測結果,每年約有20%的菌株異於往年,由此顯示稻熱病田間菌相的變化極為迅速。然而可惜的是,相關的監測研究自1995年後即已中斷,甚為可惜!所幸,近年啟動國內試驗改良場所與國際稻米研究所(IRRI)的雙向合作,使此病原菌的研究又逐漸受到重視,生理分群於是重起爐灶。根據農業試驗所陳繹年先生於2011年採集本島西部九縣市的114個稻熱病菌株,其中89菌株可對應上述七群中的六群,另有25菌株屬新的生理群,田間的主要生理群也由I、P、T群轉為K、I與新族群。以不同縣市採集的菌株種類歸納,苗栗地區可檢出六個生理群,彰化、南投和雲林地區可檢出五個生理群,臺中、臺南與屏東地區則也有四個生理群,由此顯見各地稻熱病菌相的複雜變化。 稻熱病圃檢定 除了上述對稻熱病菌的蒐集、分離外,對於水稻本身品種或品系間的病害檢定,各試驗場所亦同時進行,然而因初始檢定各單位並無統一標準,致結果難以比較。自1956年在當時的農復會補助下,陸續於宜蘭、臺中(1956~1974)或南投(1975~1981)、嘉義、屏東及臺東等五處地點設置水田式病圃,1982年後因經費之故,僅保留嘉義及關山兩處。稻熱病圃檢定主要對象為國內各試驗場所育成高級試驗以上之新品系,採兩種方式進行,一為水田式病圃,即在一般水田施以高量氮肥誘發稻熱病發生,適當時期調查葉稻熱病及穗稻熱病,目前在農業試驗所嘉義分所及臺東關山兩處進行;而另一方式則採旱田式病圃,即將欲檢定之各品種(系)稻種條播於網室內的旱田畦面,在分櫱期即可顯現抗感病情形並調查之,目前僅在嘉義分所設置。各水稻品系稻熱病圃的檢定結果,可提供各育種單位命名或推廣之參考。嘉義分所陳隆澤先生長年投身進行稻熱病圃的檢定工作,分析歷年品種(系)發現:同一品種葉稻熱病的抵抗性較穗稻熱病強,但兩者的發生存在某種程度的關連,尤其是稉稻品種有顯著的相關性,秈稻反應則略有不一致情形;而秈稻一般較稉稻的抗性較佳,但稉稻在第一期作較易罹病,但秈稻反而在第二期作罹病率較第一期作高。 抗病品種與病原生理小種的共演化關係 由多年的稻熱病圃檢定,亦可知各育種單位陸續推出抗稻熱病的新品種。然而,該抗病品種在大面積栽培之後,造成田間的生理小種菌相的演替,使得在2~4年即可能淪為感病品種。1985年由農業試驗所育成的臺農70號,為一抗葉稻熱病與穗稻熱病的品種,但在推廣四年後面積迅速擴增,卻發生嚴重的穗稻熱病,簡錦忠博士於是比較該品種在推廣前及推廣後的田間生理小種頻度,發現在推廣前可對臺農70號有致病性的菌株佔11%,而推廣後致病性菌株則提高至35%。因此可知抗病品種對於稻熱病的不同生理小種,本身也是一種選汰壓力,該品種在初始推廣種植時也許有效壓制稻熱病菌的發生,但田間終究存有可抗稻熱病的菌株,具致病能力的生理小種大量繁衍增殖,無致病力的菌株則無侵染寄主以繁殖擴增的機會,造成如上所述田間菌相的劇烈變動,也導致抗病品種臺農70號的抗性失效,旋即造成稻熱病的大發生。新的抗病品種具有壓制田間部分生理小種的能力,但數年後取而代之又有新的致病生理小種出現,育種人員又需尋找新的抗病種原,導入栽培品種中,如此不停循環的矛與盾攻防,造成特殊的作物與病原菌間的共演化關係。 抗病機制與抗病基因 作物對病原菌的抗病機制可分為兩類,一為水平抗性,一為垂直抗性。前者提供作物基礎的防禦反應,對於不同的生理小種不具明顯的專一性,因此具有較廣幅的抗病能力,但相對抗性較弱;後者則為作物的抗病基因(R gene)對應特定生理小種的非致病基因(avirulence gene),符合植物病理學家Flor(1971)提出的「基因對基因」(gene for gene)理論,可啟動後續的免疫防禦反應,具有較強的抗病性,但對於非專一對應的生理小種則無抗病能力。 稻熱病在全世界主要稻作生產區皆會發生並造成嚴重危害,兩者間的攻防關係已成為研究寄主與病原交互作用的一種模式系統,探尋稻熱病抗病基因的位置與作用亦為育成抗病品種的基礎。至今國際間已探得超過100個抗稻熱病基因,然而特別的是,該抗病基因並非平均的分佈在水稻12條染色體上,反而位於第6、11及12條染色體上特定片段呈現有多個基因群集的現象。以上發現的抗病基因已有22個被選殖完成,多數具有一般抗病基因常見的NBS-LRR (nucleotide-binding site and leucine-rich repeat,核苷酸結合位點與富含亮胺酸重複序列)保守結構;而在稻熱病菌上獲選殖的非致病基因則有13個,其中多數與水稻的抗病基因有對應關係,印證了上述的「基因對基因」理論。 國際判別品種引入 雖然國內很早即已進行稻熱病原菌的分類及病圃的檢定工作,但是病圃檢定多依賴田間自然發病狀況,各年度的病原菌生理小種會發生變化,造成各品種在多年檢定的結果有不一致的情形,且也未能清楚瞭解國內抗病品種的抗病基因究竟為何?利用已知抗病基因的判別品種在檢定時進行對照,或可推論國內品種(系)的抗病基因類型,以及各年度田間的生理小種是否改變。 近年國際稻米研究所與日本開發出具有單一遺傳背景的近似同源系,各系之間的差異主要僅在於不同的抗病基因上,包括一套在稉稻背景的LTH(麗江新團黑穀),及一套在秈稻背景的CO39。在2012年臺灣與國際稻米研究所的合作計畫中,也引入這兩套判別品種系統。適巧2013年國內第一期作雲、嘉、南地區發生較往年嚴重的稻熱病害,產量減損約10~40%,農委會下的農糧署及動植物防疫檢疫局紛紛正視此問題的嚴重性,遂討論並提出計畫,預定在各改良場轄內重要的稻熱病疫區設置簡易病圃,目的為監測易發生稻熱病疫區的生理小種演變,及各推廣品種在各地的抗感病情形。筆者也有幸參與本場在轄內新竹縣峨眉鄉的病圃檢定工作。 稻熱病生理小種之監測 各改良場自2014年起分別設置稻熱病檢定圃,主要以自IRRI引入的32個LTH近同源系,以及農糧署公告的優良水稻推廣品種為檢定對象,各品種(系)編號後依序排列,二重複,並參考國內水田式病圃的設置方式,各品種(系)種植兩行,每行種植7~10株,每隔兩個品種種植一行感病品種Lomello做為感染源,另每隔8個品種種植一行抗病品種臺農84號作為對照。筆者在新竹縣峨眉鄉所設的病圃檢定結果,發現32個品系中有11個品系在葉稻熱病的抗性表現良好,而在穗稻熱病抗性表現上則有8個品系的罹病率低於一成,顯示該批材料確實有對稻熱病抗性較佳的材料,唯需進行更長期的觀測,以期掌握各生理小種發生的變化。 分子標誌在抗稻熱病育種選拔之利用 國內有近九成的栽培面積為稉稻品種,長年育種所追求目標亦為近似的方向,造成遺傳背景窄化的現象,因此抗稻熱病基因來源受到限制;相較於此,透過與秈稻或野生稻的雜交,可望導入不同的抗病基因來源。此外,自IRRI引入的LTH或CO39近似同源系,除可作為病圃檢定用之外,因其具有已知位置的抗病基因,透過回交方式導入至國內的優良栽培品種中,並藉由分子標誌的追蹤選拔,可望加速良質米品種的育種時效,並提升抗稻熱病能力。 利用新導入抗病基因以育成新的抗病品種,或可在短期降低稻熱病的危害,但數年內恐又因病原菌生理小種頻度改變,使得新導入的抗病基因或品種失效。參考日本品種「越光」的多系育種策略,或許可為解決之道。越光為1956年育成的優良品種,具有米質優良、口感食味佳、米粒外觀晶瑩透亮等特性,至1979年躍升為日本最大栽培面積的品種,至今仍廣為農民及消費者喜愛。然而該品種對於稻熱病的抵抗力始終不佳,因此近年育種人員分別導入Pia、Pii、Pita-2、Piz、Pik、Pik-m、Piz-t及Pib等抗稻熱病基因,育成不同的越光多品系品種(Koshihikari BL)。在推廣至農民端栽培前,根據過往的生理小種監測資料,配合氣象數據以預測未來可能好發的生理小種,並依此混合數個對應的抗病品系釋出,每年所用的抗病品系及比例可能不盡相同,然此需有長年生理小種發生頻度的監測資料,以作為預測未來趨勢所需。其他品種如Sasanishiki、Nipponbare、Toyonishiki等也分別育成多系品種,且對稻熱病的抗性也有良好的控制效果。 因此,藉由多個抗病基因各別回交導入國內的優良稉稻品種中,並參考歷年稻熱病生理小種發生頻度的監測紀錄,預測未來1~3年菌相的演變,推出適當配比的多系品種,可為將來水稻育種與品種推廣的新方向。 簡禎佑 / 行政院農業委員會桃園區農業改良場助理研究員
|
|
 |
|
|
科普講座系列︰植物的光週期性 演講人︰郭華仁 / 臺灣大學農藝學系名譽教授 演講摘要 陽光是一切生命的泉源,提供了整個地球的生產者 - 植物 – 在生長各個階段的需求。植物的一生之中,從種子發芽,植株生長,至開花結果,陽光都扮演了重要的角色,本演講的內容,即在於介紹:何種波長的光會影響種仔的發芽;兩種不同的光合作用方式下,植物進行葡萄糖的合成;以及日照長短如何影響植物啟動開花的機制。地球上的植物由於演化的作用使得其受光影響的反應也不同,各種植物演化出與光不同的互動,而呈現植物多樣性的面貌。 [ 請點此下載本文全文PDF檔 ]
|
|
 |
|
|
究竟應該是分蘗、分蘖、分糱或是分櫱? 摘要 禾本科植物有一種特殊的外表性狀(trait),農藝學上稱為分櫱,也就是說每一植株除了主要的莖桿之外,還有側生的莖桿,類似古代人倫關係中的嫡子與庶子,此處的庶子亦稱為孽子。「孽」的上半部是「薛」字下半部是「子」字,將「子」字以「木」字取代即為櫱的俗體字「蘖」。由於櫱字是罕用字,經常以俗體字的形式「蘖」出現。此俗體字「蘖」的外型恰巧與「蘗」十分相似,因此「分蘖」一詞在農藝學相關的文獻上普遍書寫成或是印刷成「分蘗」。農藝領域的專家學者,習而不察,但是隨著科普教育的發展,農作物的知識逐漸普及,農藝領域之外的社會大眾很困惑,為何「分蘗」唸成「ㄈㄣㄋㄧㄝ‵」而不是「ㄈㄣㄅㄛ‵」。本文嘗試去瞭解造成此種混淆的可能原因,並追溯此混淆究竟從何時開始的,並希望推廣正確的使用。 [ 請點此下載本文全文PDF檔 ] 彭雲明 / 磯 永吉學會秘書長
|
|
 |
|
|
秘書組 預定在 2016/3/25 召開理監事會議(第二屆第三次理監事會議), 審查案: 人事異動同意案: 文史與出版委員會 2016/03/01 與黃臺香(風景文化事業發行人)商談《米報文集2013-2015》的排版與印刷 北部蓬萊米走廊聯盟委員會 2015/12/23 竹子湖原種田故事館舉行開幕式 志工事務委員會 2016/01/12 志工幹部會議(1月份) 作物科學科普講座委員會 2016/12/10 邀請林華慶博士(臺博館副館長)演講—推行友善農業成功的案例分享 古蹟建物與文物委員會 2016/02/28 磯小屋建築藍圖重新製作完畢,以畫架方式展示
|
|
|
|
|
| 磯小屋米報著作權屬於國立臺灣大學磯永吉學會br> 臺北市羅斯福路四段一號 TEL:(02)33664754 |  |